370-371 


Гормони. гідрофільні гормони
Метаболізм пептидних гормонів
 А. Біосинтез
А. Біосинтез
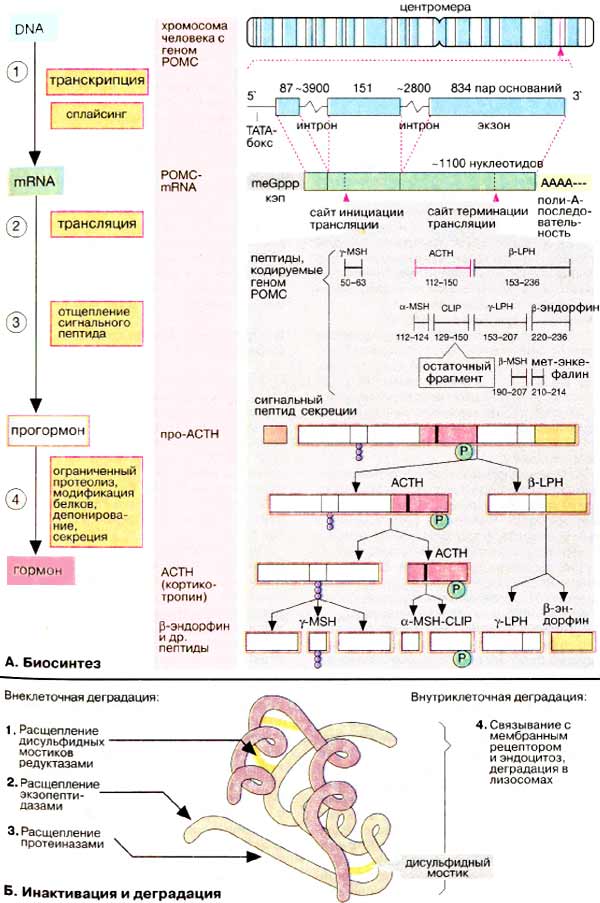
На відміну від стероїдів пептидні і білкові гормони є первинними продуктами біосинтезу. Відповідна інформація зчитується з ДНК (DNA) на стадії транскрипції (див. С. 240 ), А синтезована гяРНК (hnRNA) звільняється від інтронів за рахунок сплайсингу (1). мРНК (mRNA) кодує послідовність пептиду, який найчастіше істотно перевищує за молекулярною масою зрілий гормон. Вихідна аминокислотная ланцюг включає сигнальний пептид і пропептид - попередник гормону. Трансляція мРНК відбувається на рибосомах за звичайною схемою (див. С. 246 і сл.) (2). Спочатку синтезується сигнальний пептид. Його функція полягає в тому, щоб зв'язати рибосоми на шорсткою ЕПР [Шер (rER)] і направити зростаючу пептидную ланцюг в просвіт Шер (3). Синтезований продукт є попередником гормону, прогормоном. Дозрівання гормону відбувається шляхом обмеженого протеолізу і подальшої (посттрансляционной) модифікації, наприклад утворення дисульфідних містків, гликозилирования і фосфорилювання (див. С. 226 ) (4). Зрілий гормон депонується в клітинних везикулах, звідки секретується в міру необхідності за рахунок екзоцитозу.
Біосинтез пептидних і білкових гормонів і їх секреція знаходяться під контролем ієрархічної системи гормональної регуляції (див. С. 360 ). У цій системі в якості вторинного месенджера беруть участь іони кальцію; збільшення концентрації кальцію стимулює синтез і секрецію гормонів.
Аналіз гормональних генів показує, що іноді багато абсолютно різні пептиди і білки кодуються одним і тим же геном. Одним з найбільш вивчених є ген проопиомеланокортина [ПОМК (POMC)]. Поряд з нуклеотидної послідовністю, відповідної кортикотропіном [адренокортикотропний гормон, АКТГ (АСТН)], цей ген включає перекриваються послідовності, що кодують ряд невеликих пептидних гормонів, а саме α-, β- і γ- ме ланотропінов [МСГ (MSH)], β- і γ- ліпотро пинов (ЛПГ (LPH)], β- ендорфіну і мет-Енке Фаліна (див. рис. 343 ). Останній гормон може також утворюватися з β-ендорфіну. Прогормоном для цього сімейства є так званий поліпротеїнів. Сигнал про те, який пептид повинен бути отриманий і секретувати, надходить із системи регуляції після завершення синтезу препропептіда. Найбільш важливим секретується продуктом, отриманим з гипофизарного поліпротеїну кодованого геном ПОМК, є гормон кортикотропін (АКТГ), який стимулює секрецію кортизолу корою наднирників. Біологічні функції інших пептидів до кінця не з'ясовані.
Б. Інактивація і деградація
Деградація пептидних гормонів часто починається вже в крові або на стінках кровоносних судин, особливо інтенсивно цей процес йде в нирках. Деякі пептиди, що містять дисульфідні містки, наприклад інсулін, можуть инактивироваться за рахунок відновлення залишків цистину (1), Інші білково-пептидні гормони гідролізуються протеиназами, а саме екзо (2) (по кінцях ланцюга) і ендопептидаз (3). Протеолиз призводить до утворення безлічі фрагментів, деякі з яких можуть проявляти біологічну активність. Багато білково-пептидні гормони видаляються з системи циркуляції за рахунок зв'язування з мембранним рецептором (див. С. 372 ) І подальшого ендоцитозу гормон-рецепторного комплексу. Деградація таких комплексів відбувається в лізосомах, кінцевим продуктом деградації є амінокислоти, які знову використовуються в якості субстратів в анаболічних і катаболічних процесів.
додаткова інформація
Ліпофільні і гідрофільні гормони мають різний напівперіод існування в системі циркуляції (точніше біохімічний напівперіод, t1 / 2). У порівнянні з гідрофільними гормонами (t1 / 2 кілька хвилин або годин) ліпофільні гормони живуть значно довше (t1 / 2 становить кілька годин або днів). Біохімічний напівперіод гормонів залежить від активності системи деградації. Вплив на систему деградації лікарськими препаратами або пошкодження тканин може викликати зміну швидкості розпаду, а отже, і концентрації гормонів.